 |
|
|
|
|
|
|
|
|
3.3. Die Wachstumsenergie |
|
|
|
|
|
um wichtigsten Wohl und W des biologischen Lebens gehören Wärme und Wasser. Das Wachstum wird über Oberflächen, vor allem Membranen, durch Wasseraustausch mit verschiedenen Wasserqualitäten gewährleistet. Der Wasserentzug ist am besten bei Landpflanzen in einfachen wägbaren Lysimetern zu quantifizieren. So soll dieses Kapitel ein Hohelied auf die wärmegetriebene Transpiration werden. Wir führen den Todesstoß gegen den Irrglauben, daß die Photosynthese ein DIREKT photonengetriebener Sonderprozeß ist.
Wärme kann entgegen dem zweiten Hauptsatz an Oberflächen gesammelt und in strukturierte Oberflächenspannung umgewandelt werden. Sie ist die Grundlage des gesamten Lebens. Am Ende des Lebens muß aber alles wieder zurückgegeben werden. Ohne Opferungssterben geht kein Wachstum, was dem Leben identisch ist.Wärme ist die URENERGIE. Alle Energie wird mit Leichtigkeit in Wärme umgewandelt. Bei jedem Prozeß entsteht Wärme. Sie ist nicht gequantelt, wie sie in den Schulphysiklehrbüchern kaum nachlesen können. Die wichtigste Energie nicht gequantelt!!!!!!????? Der umgekehrte Prozeß des Wärmesammelns und -wandelns kostet Organisationstalent und Intelligenz, über Oberflächen diese Wärme wieder einzufangen und zu konzentrieren. Das Einfangen geht isotherm auf breiter Basis, das Abgeben adiabatisch und konzentriert, sehr schnell und ohne Energieaustausch mit der Umgebung vor sich. |
|
|
|
|
|
|
|
|
|
|
|
Ich habe das einmal den ADIABATISCHEN TRICHTER genannt und gehe abstrakt-konkret im Anhang darauf ein.
Nun zur Photosynthese als repraesentatio mundi und historii. Auch bei diesem Prozeß muß gemäß biogenetischem Grundgesetz das Licht in Schwingungsprozesse, Wärme, umgewandelt werden. Denken Sie an die theoretische Ableitung der Molenwärme von Einstein, Debye und Raman. In der Biologie geschieht das konkret über die Umwandlung der Wärme in Oberflächenspannung, deren Energie über Mikrokugelexplosionen freigesetzt und in maßgeschneiderte Stoffwechselenergie gewandelt wird. Obwohl einige Experimente reichen würden, um meine Interpretation zu illustrieren und wärmstens zu unterstützen, werde ich trotzdem Beispiele in großer Zahl anbieten, um auch den letzten Zweifel auszuräumen, wenn auch nicht den aller- letzten.
Es soll ja heute noch Leute geben, die glauben, daß die Welt eine Scheibe ist. Sehen heißt glauben und nicht wissen. Diese können nicht über ihren Tellerrand hinaus und haben so für ihre unmittelbare Umgebung sogar recht, wenn sie nicht auf die ganze Erde extrapolieren würden.
Wärme und Wasser machen die Erträge und nicht in erster Linie das Licht, das ebenfalls erst in Wärme gewandelt werden muß.
Die Energie eines Rotquanten reicht nicht aus, ein T-Wassermolekül zu spalten. Nach dem photochemischen Äquivalenzgesetz sind aber Mehrphotonenprozesse verboten. Sie sind wegen der statistischen Natur des Lichtes unmöglich. Außerdem steht in jedem Lehrbuch der Physik, daß allein schon durch die Wärmestöße das Wassermolekül in Ionen verwandelt wird, rein spontan. Ionen sind natürlich sehr reaktionsfreudig, besonders das aggressive PROTON.Der Ertrag wird in erster Linie von Wärme und Wasserangebot bestimmt. Nehmen wir zuerst eine instruktive Arbeit (44), die zeigt, daß mit zwei Kurven, von denen die eine die Abhängigkeit des Ertrages vom Niederschlag und die andere von der Temperatursumme darstellt, die gesamte Stoffproduktion aller Landpflanzen genau geschätzt werden kann. Die Autoren gehen dabei nur von der einen aus, deren Angebot im Mangel ist!!! Im hohen Norden ist das bestimmt die Temperatursumme, in der Wüste die Niederschlagsmenge. Das Licht spielt nur indirekt eine Rolle, weil die Sonneneinstrahlung die Temperatur im gewissen Maße bestimmt. Die Anpassung an die tatsächlich gemessene Stoffproduktion ist erstaunlich. Als nächstes sollen hier die Rekordertragspflanzen aus verschiedenen Biotopen genannt werden und die Hektarerträge an TROCKENmasse. Je feuchter und wärmer das Milieu, um so höher die Trockenmasse pro Hektar und Jahr. |
|
|
|
|
|
|
92t/ha*a
|
Algen in Israel (Chlorophyceae)
|
|
67t/ha*a
|
C-4-Pflanzen (Zuckerrohr-Hawaii)
|
|
22t/ha*a
|
C-3-Pflanzen (Zuckerrübe-Holland)
|
|
60t/ha*a
|
Gewächshaus (Tomate-Holland)
|
|
|
|
|
|
|
Dazu folgende Bemerkungen. Im Wasser ist das Licht geringer als auf dem Lande. Im Gewächshaus sind nur 50-70 Prozent des Lichtes wie im Freiland. Freilandtomaten haben einen wesentlich geringeren Ertrag, natürlich wegen der fehlenden Temperatur. Die Diskrepanz zwischen jetziger Theorie und Praxis wird aus der Tatsache abgeleitet, daß den Physikern und damit auch den gläubigen Botanikern die Wasseroberflächen mit ihren großen Energien entgangen sind, die besonders im warmen Wasser stecken, weil hier eine Oberfläche besonders schnell wieder aus der Umgebungswärme aufgebaut werden kann.
Die jetzige Photosynthesetheorie geht mehr vom Zählen aus. Die hereinkommenden Quanten werden gezählt und ein Bruchteil der Energie bei jeder Zählung genutzt. Meine Theorie geht mehr von der Wägung aus. Alle Quanten, besonders natürlich die des sichtbaren Lichtes, weil die infraroten von den Blättern wegen der Hydrophobie des Protoplasmas zurückgestrahlt oder durchgelassen werden, werden mit einem sehr kleinen Maß gewogen, in Einheitsquanten gesammelt und nach Maß wieder in größere umgewandelt. Die Photosynthese wäre dann eine Art Umkehr des superschwachen Leuchtens.
Als Kompromiß könnte man sagen, daß die Pflanze mittels Wägung die Wärmequanten, was das auch immer sein mag, zählt. Auch in der Technik werden die Stückzahlen von identischen Schrauben mittels Waage gezählt. Keine der beiden Theorien ist von vornherein von der Hand zu weisen. Beide sind irgendwie wahr, aber meine ist wahrer.
Nehmen Sie die zahlreichen Experimente mit reinem blauen oder rotem Licht, die beide in der gleichen Photonenzahl pro Quadratmeter und Zeiteinheit angeboten wurden. Die Blaupflanzen hatten einen fast doppelt so hohen Trockenmasseertrag wie die Roten. Ganz genau dem Energiegehalt pro Quant entsprechend. Das ist beste Wägetheorie. So viele Experimente bei so vielen Pflanzenarten gibt es bis jetzt, aber keiner kommt auf den adiabatischen (Nürnberger) Trichter. Die Strahlungsmessung in EINSTEIN ist physiologischer Unsinn. Ich als Biologe verlange Selbstbewußtsein vor den physiologisch unbedarften Physikern. Das Licht ist nicht nur Quant.
Ich kann sehr einfache Ertragsmodelle machen, ähnlich wie (44), die nur einen Faktor der quantitativen Physiologie berücksichtigen. Die beste Schätzung kommt bei alleiniger Einbeziehung der Transpiration heraus, obwohl dieser Prozeß auch noch die unproduktive Verdunstung beinhaltet, weil ein Lysimeter beide Prozesse, die aktive und passive Wasserabgabe, nicht unterscheiden kann. Es gibt für jede Pflanzenart andere Transpirations-Ertragsbeziehungen. Alle Pflanzen haben verschieden gute Techniken entwickelt, die den Grundprozeß variieren.
In Holland wurde folgender Versuch gemacht. In einem Gewächshaus wurde das Licht gedrosselt, aber durch klimatische Veränderungen die Transpiration trotzdem erhöht. In dem Vergleichshaus wurde mehr Licht gegeben aber durch das Klima die Transpiration gedrosselt. Der Ertrag bei weniger Licht war signifikant höher. Was mich hier besonders erbost war die Tatsache, daß ich bei diesen Damen und Herren in Holland war und meine Vorstellungen anpries. Sie haben sogar meinen Nachfolger in Großbeeren, der jetzt mein Ertragsmodell in Großbeeren weiterbehandelt, zu einem dreimonatigen Studienaufenthalt eingeladen, um mein Ertragsmodell besser kennenzulernen, aber meine Grundvorstellungen lehnten sie mit dem Hinweis ab, daß alle Stoffe, die im Photosynthesezyklus vorkommen, doch nachgewiesen seien. Quasi gesehen wurden. Als wenn das ein Gegenbeweis wäre. Sehen und Interpretieren wird als identisch angesehen. Es muß doch an unserer Fernsehgesellschaft liegen, wie Neil Postman sagt.
Die Photosynthese wurde nach plötzlichem Abschalten des Lichtes im Dunkeln weitergemessen (48). Bei allen untersuchten Pflanzen nahm die CO2 - Verarbeitungsgeschwindigkeit erst nach Stunden den Wert Null an. Je höher letztere am Tage war, um so kürzer war der Weg zu Null, weil die Opferungsflußanspannung bei viel Photosynthese natürlich größer ist, weil relativ mehr Elementararbeiter am Werke sind und weniger Erholungszeit zur Verfügung haben. Wer treibt die Photosynthese im Dunkeln so viele Stunden an? Die Antwort können Sie wohl schon selber geben.
Wenn Gurkenpflanzen im September gesetzt werden, so haben sie am Anfang einen Energieüberschuß von der Sonne. Wenn der Dezember kommt, ist die Pflanze sehr groß und hat eine stärkere Atmung als Photosynthese, geht aber nicht ein und stabilisiert sogar die Trocken- und Frischmasse. Woher kommt die Energie?
Die Atmung wurde an Gurkenpflanzen in Abhängigkeit von der Temperatur gemessen. Sie nimmt bei 5 K Temperaturerhöhung um das zwei- bis dreifache zu. Bei Pflanzen, die bei 24 Grad Celsius im Gewächshaus gehalten werden, dürfte somit kein Ertrag mehr zustande kommen, was aber bei weitem nicht der Fall ist. Der erste Ertrag kommt sogar eher als bei kälter gehaltenen Pflanzen. Der Gesamtertrag am Ende des Jahres ist aber natürlich wesentlich niedriger als bei den letzteren. Woher kommt die meßtechnisch nicht erfaßte Energie? Nur aus der Gewächshauswärme.
Im Herbst, wenn es ordentlich warm wird wider jeden Erwartens, blühen plötzlich alle Bäume trotz eklatanten Lichtmangels. Denken Sie auch an die Gewächshauspflanzen im Vergleich zu ihren Freilandgeschwistern.
Das häufigste und wichtigste Ferment ist das RUBISCO. Es bindet das CO2 an eine drei C-Atome lange Kette. Es braucht viel Energie, weil das Kohlendioxid ein anerkannt giftiges Produkt ist. Es zu verarbeiten ist Schwerstarbeit. Über die Gase, die die lebenswichtigen Kapillaren verstopfen können, haben wir schon einiges gehört. Das RUBISCO erlaubt sich darüber hinaus noch die Unerhörtheit, daß es gleichzeitig die Veratmung, also den Zerfall der soeben aufgebauten Ketten, katalysiert. Das nennt sich Lichtatmung. Bis jetzt vollkommen unverständlich, aber in dem Augenblick nicht mehr, wenn man weiß, daß wärmegetriebene Opferungsflußenergie oder Eustreß gebraucht wird, um zu wachsen und auch die Fermente, wie den Weltmeister RUBISCO, der das in eigene Regie übernommen hat, zu renaturieren oder neu aufzubauen.
Aber außer der Wärme wird auch noch Kohlendioxid geliefert, auf das kein Lebewesen verzichten kann. Bei Mikroorganismen wurde das zuerst festgestellt. Sie können nur im Verband gedeihen, wenn Ihnen das Kohlendioxid entzogen wird., weil das durch die Atmung entstehende Gas natürlich bei vielen nicht so schnell wegdiffundieren kann. Einzelzellen gehen ein. Die haben eine zu große relative Berührungsfläche mit der Außenwelt. An den Photosynthesezentren wird sehr viel CO2 verbraucht, so daß es zu einer lebensbedrohlich geringen Konzentration kommen kann.
Bei C4-Pflanzen gibt es praktisch keine Lichtatmung, weil hier das lebensnotwendige CO2 durch die Tätigkeit der PEP-Carboxylase geliefert wird, die das Kohlendioxid an Malat oder Aspartat bindet und das Kohlendioxid mittels organischer Säuren aktiv transportiert, so daß an den kohlendioxidverbrauchenden Photosynthesezentren immer genügend lebensnotwendiges Gas ist.
Warum verzichten die C3-Pflanzen auf den PEPC-Prozeß? Er läuft nur im Warmen gut. Er verlangt einen langen und teuren Weg. Die mehr passive Kohlendioxidpermeation der C3 - Pflanzen ist dagegen praktisch nicht von der Temperatur abhängig. Sie geht nahezu automatisch vor sich.
Auch die Hillreaktion geht mit Kohlendioxid viel besser, was von den Vertretern der jetzigen Theorie gern verschwiegen wird. Bei der Photosynthese wird wahrscheinlich immer der Bicarbonatkomplex H+...HCO3 - gespalten.
Wahrscheinlich spielt der Carbonatkomplex noch eine wesentliche Rolle bei der Ausfällung der lebensnotwendigen Calcium- und Magnesiumionen, die bei der Erregung in die hydrophilen Poren eindringen. Siehe das Kapitel über die Membranen und die temporären Poren. Durch die Ausfällung wird den beiden wasserhüllenreichen Erdalkaliionen die ungemein lebensnotwendige H+...OH-Ionenhülle entrissen. Praktisch alle Kalkgebirge sind durch biologische Tätigkeit entstanden. Denken Sie an Muscheln, Schnecken , Eierschalen usw. Auch an das Phosphat sei in diesem Zusammenhang erinnert, besonders das welches beim Zerfall des ATP entsteht. Phosphate sind im allgemeinen schwerlösliche Verbindungen, die ebenfalls wie das Carbonat Calcium aber auch das genauso wichtige Magnesium fällen können, um an das lebendige Wasser, das H+...OH-, heranzukommen. ATP kann so bei vielen Prozessen den Auslöser spielen, quasi wie ein Hormon wirken.
Kommen wir zur sogenannten "idle" respiration (49). Bei Pflanzen gibt es zwei Atmungswege. Die erste, die Cytochromatmung, erzeugt vor allem ATP und kaum Wärme. Die "idle" erzeugt nur im ersten Schritt ATP und dann "nur" Wärme. Angeblich, um "überflüssige" Kohlehydrate abzubauen, die mit der Wärme verpuffen. In den Teilen der Getreidepflanzen, die am schnellsten wachsen und die meisten Kohlehydrate umsetzen, in den Samen, Früchten oder Karyopsen, ist die "idle" aber am größten (50). Wo sind da die überflüssigen Kohlehydrate? Das Getreide stößt einen Großteil der Fruchtansätze ab, um so mehr, je schlechter es ihm geht. Die Kohlehydrataufnahmefähigkeit ist immer groß genug.
Die wärmegetriebene Wasserpumpe wurde bei Kohlrabi direkt nachgewiesen (51). Die Pflanzen wurden mit so wenig Licht versorgt, daß ihre Photosynthesebilanz negativ war. Die Trockenmasse nahm ab, die Frischmasse, besonders die der Knolle, nahm aber um ein erkleckliches zu. Kommentar überflüssig. Auch bei diesem Menschen, er wohnt und lehrt in Hannover im goldenen Westen, war ich noch heimlich vor der Wende gewesen. Er sagte mir, daß er mein Ertragsmodell viel besser als sein eigenes fände, aber meine Grundprämissen wollte er nicht anerkennen. Trotz seines Versuches. Er ist zumindest sehr standhaft.
Ein noch drastischerer Versuch wurde bei Bohnen- epidermis durchgeführt. Die Photosynthese wurde mit Tentoxin schön spezifisch und total ausgeschaltet. Die Epidermis wurde entweder nur mit Weißlicht bestrahlt oder nur mit Wachstumshormonen behandelt. Beide Male nahm die Masse der Epidermis zu!! Kommentar überflüssig!!
Auch mit diesem Herrn (52) habe ich gesprochen. Er stritt meine Ergebnisse mit dem gasbefreiten Wasser einfach ab und verlangte von mir, das ich nachwiese, daß das Wasser wirklich in die Zellen eindringe!!! Wie kann sonst ein Turgor zustandekommen, wenn nicht das Wasser in die Zelle eindringt??? Er und seine Kollegen lehnten später ein Gespräch mit mir ab, weil sie mich wahrscheinlich für bekloppt hielten.
Auf einer ähnlichen Ebene liegen die Ergebnisse von (55). Die Autoren arbeiteten ebenfalls mit Epidermis und wiesen nach, daß C14O2 ebenfalls im Dunkeln fixiert wird. Daß es mit Licht viel besser geht, steht nicht zur Debatte. Es geht um das Prinzipielle.
Wir erinnern an die Membran-ATPasen, die mit schonenden nicht denaturierend wirkenden oberflächenaktiven Stoffen behandelt wurden und ihre Intensität vervierfachten. Auch hier war der Herr, der mit der Bohnenepidermis, beteiligt. Er stellt in dem Artikel die gleichen Fragen nach dem warum, aber an mich hat er nie wieder gedacht.
Was macht aber nun das ATP? Es ist nur in der lebenden Zelle wirksam, weil es hydrophob ist und im Wasser hydrolysiert wird. Es gibt Vitalitätstests von Bakteriensuspensionen, die auf der Bestimmung der ATP-wirk-samkeit beruhen, weil die ATP-Aktivität mit dem Leben erlischt. Einen Schlüssel zu der Beantwortung dieser Frage könnte die sogenannte polyadenylierte RNS (56) geben und die Tatsache, daß in jungen wachsenden Zellen von Bakterien 1...6mg ATP pro Gramm Biomasse aber in ausgewachsenen nur 0.05-0.8mg/g gemessen wurden.
ATP ist vor allem für die Renaturation und den Neuaufbau der Eiweiße mittels der RNS zuständig. Da ein ständiger Opferungsfluß notwendig ist, je mehr Wachstum um so mehr, kann bei keiner Fermentarbeit auf das ATP verzichtet werden, weil es mit ATP besonders gut geht, nicht weil es prinzipiell dabei sein muß. Phosphate beeinflussen die Oberflächenspannung des Wassers besonders stark. Auch Aktinfasern können sich ohne ATP außerhalb des lebenden Zellverbandes zusammenrollen, aber mit ATP und Myosin in der lebenden Zelle ungleich schneller und wirksamer.
Ich will es einmal so formulieren. Prinzipiell ist ATP überflüssig, aber im Hinblick auf die harte Konkurrenz der Nachbarn auf keinen Fall.
Über die Rolle des ATP war ich mir jahrzehntelang nicht im klaren. Es ist mir erst 1992, sechs Jahre nach Geburt meiner Wärmewasserwellentheorie, gelungen, zu einem einigermaßen befriedigenden Ergebnis, zu einer Verallgemeinerung aller mir bekannten Eigenschaften in einem Wort, zu kommen.
ATP spielt in der lebenden Zelle ohne Zweifel eine Schlüsselrolle. Es ist bei allen metabolischen Prozessen irgendwie dabei, besonders wenn es um den Um- und Neuaufbau in der jungen Zelle geht. Es gilt nach der jetzigen Theorie als der Hauptenergieträger und - überträger.
Ich bin dagegen, weil das ATP viel zu wenig in der Zelle vorhanden ist, besonders wenn man die obigen Zahlenwerte mal auf die Frischmasse umrechnet. Es wurde einmal ungefähr ausgerechnet, wieviel Energieumsatz der menschliche Körper an einem Tag hat und wieviel kg ATP vorhanden sein müßten, wenn es nur einmal am Tage in Funktion treten dürfte. Man kam so ungefähr auf 70kg ATP. Wer lügt zahlt einen Taler. Die Autoren weisen somit auf das Wunder Leben, das angeblich eine so große Umschalthäufigkeit des ATP verwirklichen kann. Das würde vielleicht noch angehen, aber wenn Sie dann noch an die verblüffend hohe Beweglichkeit des Stoffes denken, weil überall in der Zelle ständig Energieumsetzungen stattfinden. Denken Sie besonders an die Bakterienzelle, die einen hundertmal höheren Stoffwechsel pro Masseeinheit hat als die der Mehrzeller. Das ist einfach zuviel der Wunder. So schnelle Moleküle gibt es einfach nicht. Ich habe zwar den Geist in der Natur und das Wunder des Lebens beschworen, aber den mathematisch faßbaren im unendlichen Limes. Das ist nichts esoterisches, wie mir doch tatsächlich einige weismachen wollen. Sie führen deshalb lauter fadenscheinige Gründe vor, um dieses Buch nicht zu drucken. Sie haben fast alle einen Horror vor der Mathematik, können und werden das aber niemals zugeben.
Kommen wir zu den entscheidenden Eigenschaften des ATP, aus denen Sie eigentlich schon selbst auf das richtige Wort für das ATP kommen müßten.
1. Sehr gering konzentriert
2. stark hydrophob
3. Wird bei der Stoffwechseltätigkeit schnell und leicht zu ADP und dem reaktionsfreudigen Phosphation abgebaut, welches die L-"Wasser"reichen Calcium und Magnesiumionen fällt, was weiter oben schon im Hinblick auf das lebensnotwendige Kohlendioxid und seine ionogenen Derivate erwähnt wurde.
4. Es unterliegt der Bioakkumulation und hat trotz seiner geringen Konzentration solch hohe Wirksamkeit.Richtig kombiniert.
Es hat genau die Eigenschaften eines HORMONS.
Es liefert Nachrichten über die Großwetterlage in der inneren Zelle. Es ist aber ein Bote mit einer starken Peitsche in der Hand, aber kein Befehlshaber, sondern wie ein Aufseher in der Sklaverei. Ein Aufseher ist bekanntlich mehr wert als zehn Arbeiter. Ich mache da keinen Spaß, denn die Zelle versteht keinen und weiß was so ein ATP-Aufseher wert ist. Er löst mit wenig aber konzentrierter Energie an den schmerzhaftesten Stellen der Proteine oder des Protoplasmas die Energien, die dort schlummern, aus. Dabei liefert es noch zusätzlich H+... OH-. Zuckerbrot oder Peitsche???
ATP ist ein Kürzestweghormon. Im Gegensatz zum Adrenalin oder den Geschlechtshormonen. Als Zwischenstufe gibt es die Gewebshormone. ATP ist wirklich prinzipiell nicht notwendig. Denken Sie noch einmal an die Exoenzyme, wobei die sogenannten Weißfäulepilze, wie der Austernseitling, welche abgeben, die sogar Lignin knacken, was der Mensch erst in der Neuzeit lernte. Auch die Geißeln der Bakterien, die haargenau wie ein Flugzeugpropeller arbeiten, kommen ohne ATP aus (80). ATP wurde erst auf einem relativ hohen Entwicklungsstand erfunden, als die Zellen eine bestimmte Größe überschritten und die mikroheterogenen Bezirke koordiniert zur Paranekrose geführt werden mußten.
Der Clou vont Janze sind die schon erwähnten Methanbakterien. Der Autor schreibt sogar ausdrücklich, daß zwei Wassermoleküle in einer unerfindlichen Form an der Reaktion teilnähmen. Er wüßte nicht warum. Als Arbeiter natürlich, aber Wasser ist eben schon verbrannt. Da kann keine Stoffwechselenergie mehr herkommen. Haha, kann ich da nur sagen.
Auch die berühmte Summenformel der Photosynthese zeigt, daß hierbei die Entropie abnimmt. Wer übernimmt hierbei den Entropieexport, um in der Sprache der Herren Eigen und Prigogine zu sprechen (54)???
Einen noch schöneren Versuch machte eine deutsche Truppe mit Samenkeimung. Sie ließ einen Lichtkeimer im Dunkeln keimen und behandelte ihn alle 24h eine Stunde lang mit 40 Grad Celsius. Dieser Keim sah danach genauso aus, als wenn er im Licht gekeimt wäre. Licht verwandelt sich im Pflanzenkörper in Temperatur (53).
Welche Rolle spielt also das Licht? Es wirkt bei der Photosynthese weit besser als die direkte Wärme von außen, weil es in die Zellen bis an alle Photosynthesezentren sehr schnell herankommt als Strahlung. Es versorgt die INNEREN Oberflächen mit Energie. Die nicht photosynthesierenden Teile sind durchsichtig.
Das Licht ist notwendig aber nicht ausreichend. Gurkenblätter sterben bei Licht und 3 Grad Celsius sehr viel schneller ab als bei gleicher Temperatur im Dunkeln. Um das noch deutlicher zu machen, betrachten wir einmal Alpha- und Betastrahler in ihrer Gefährlichkeit für den Organismus. Erstere Strahlen haben eine so geringe Durchdringungsfähigkeit, daß sie als ungefährlich gel-ten. Sie durchdringen nicht einmal Papier, geschweige denn Kleidung oder die obersten Schichten der Haut. Betastrahlen haben eine weit höhere Durchdringungsfähigkeit und gelten nicht umsonst als gefährlicher. Wenn wir aber den Alphastrahler in den Körper aufnehmen, wird er plötzlich viel gefährlicher, denn ALLE Strahlen wirken nun ionisierend und damit tödlich für die betroffenen Moleküle. Der Betastrahler schießt auch im Körper viele Elektronen nach außen, so daß nur ein Teil im Körper ionisierend wirkt. Am gefährlichsten im Innern sind thermische (WÄRME) Neutronen. Sie prallen an den Protonen, den Wasserstoffkernen der biologischen Substanz (L"Wasser"), ab und überlassen so ihre gesamte Todesenergie dem Körper.
Analog ist das mit der Lichtstrahlung. Von außen ist sie wesentlich wirksamer, weil sie wie der Betastrahl in den durchsichtigen Pflanzenkörper eindringt. Von innen ist Wärme wesentlich wirksamer, weil sie Allkontakt bedeutet, was zu einer Schwingungbeschleunigung (Kraft) führt (Wärme, Wasser und WELLEN).
Der angebliche Unsinn, das Licht nicht direkt wirken zu lassen, wandelt sich in tiefe Weisheit. Durch Umweg geradewegs ins Ziel der optimalen Stoffproduktion oder Erhaltung, des optimalen Wirksamwerdens der kostbaren (Licht-) Energie.
Bevor wir noch ganz konkret auf die vollendete Transpiration der Stomataschließ- und der Nebenzellen kom-men, wollen wir es nicht versäumen auch die Tiere, speziell die Säuger mit ihrer Schwitztätigkeit zu beachten, die angeblich der Abkühlung dient. Bei Anstrengung fließt der Schweiß in Strömen herunter. Die Pflanzen sind also viel ökonomischer, auch wenn die Transpiration nur als Kühlung dienen sollte. Eine Ausnahme sind vielleicht die Weltmeister an Ausdauer unter den Säugern, die Caniden, die keine Schweißdrüsen besitzen und das Hecheln zur Vollendung getrieben haben. Fast schon eine pflanzliche Transpiration mit ihrem Energiegewinn?? Aber wohl nicht, denn den Hunden fließt, besonders bei Anspannung, auch der Speichel ziemlich flüssig aus dem Maul. Schwitzen dient kaum der Abkühlung. Denken Sie an die Tuaregs in der Sahara mit ihrer schwarzen Kleidung, trainierende Sportler, Soldaten in Gummikleidung und den Schnuffis. Alle verhindern die Transpiration, aber nicht das Schwitzen. Da kühlt nichts mehr. Jedem Sportler ist es streng verboten, nackt zu trainieren, weil durch Transpiration die Wärmeverluste zu groß wären.
Wenn die eigentliche Transpiration, die Gaswerdung und Verdunstung, verhindert wird, der eigentlich abkühlend wirkende Vorgang, stirbt kein Mensch. Wenn das Schwitzen verhindert wird, stirbt man sehr schnell. Auch in der kalten Nacht und im Wasser. Bei Ausfall der Nierenfunktion stirbt man auch, aber lange nicht so schnell. Das Verhindern des Schwitzens wirkt kaum durch Vergiftung des Körpers. Schwitzen ist Leben.
Am Schluß diese Exkurses, der Sie hoffentlich ins Schwitzen gebracht hat, erinnern wir an die Weltrekord-, Eis-, Kanal- u.a. Schwimmer, die ebenfalls große Mengen an FRISCHmasse durch die Rippen schwitzen, obwohl sie das gar nicht dürften. Abkühlung ist genug da. Ich hoffe, daß alle meine Gegner nun gebührend ins Schwimmen und damit auch Schwitzen geraten sind.
Alle Wassertiere und -pflanzen haben einen regen Oberflächenwasseraustausch. Denken Sie an die leistungsfähigsten Schwimmer, die Delphine und Wale, die an Land sterben, weil ihre Haut austrocknet, die im Wasser wahrscheinlich durch Transpiration große Energiemengen gewinnen kann, so daß der Delphin viel schneller schwimmt als seine zuverlässig gemessene Muskelkraft eigentlich hergibt. Er setzt wie ein Campherboot die Oberflächenspannung stark herab und bekommt somit einen Extraenergieschub beim Schwimmen.
Betrachten wir die Wasserabgabe eines Nelkenbestandes von einem Quadratmeter Fläche. Er überführt im Jahr 850000cm3 Wasser von der flüssigen in die gasförmige Phase. Das entspricht einer Verdampfungsenthalpie von rund 2*106J. Die Globalstrahlung in diesem Gewächshaus betrug 1.74*106J. Davon nimmt die Pflanze nur einen Teil auf, weil sie ja wie schon bekannt das infrarote zurückstrahlt oder durchläßt. Außerdem sind zwischen den Pflanzen immer Lücken, so das ein Teil der Strahlung in den Boden geht und in Wärme verwandelt wird. Die Strahlungsenergie, die von der Pflanze aufgenommen wird, reicht also bei weitem nicht aus, um die Tran-spiration zu betreiben. Die Wärmeenergie muß also ran. Ich zitiere hier noch einige Arbeiten, die das alle belegen, wenn Sie sich in einigen der Mühe unterziehen, die Bilanz selber aufzustellen (57-62).
So weit so klar. Ich bin nicht der erste, der auf diese Diskrepanz aufmerksam wurde. Die entscheidende Frage, die noch von keinem Autor gestellt wurde, ist die, welche Art von Energie für die Photosynthese, die ja ein quantengetriebener Prozeß sein soll, übrig ist??? Es bleibt nur der Schluß, daß die Photosynthese im Wasserstrom der Transpiration mitgetrieben wird. Die Transpiration beträgt auch in der Nacht noch ein Viertel bis ein Drittel der am Tage, wenn Temperatur und Luftfeuchte konstant gehalten werden, was in Küvetten gut möglich ist. Nachts wird die Photosynthese nur langsam abgestellt, wie wir schon erfahren haben. Wir müssen hier noch einmal betonen, daß es mit dem durchdringenden Licht viel besser geht.
Denken Sie noch einmal an den Emersoneffekt. Es wird immer viel zuwenig herausgestellt, daß die Wasserspaltung mit zwei Lichtpaketen sich im Vergleich mit einem Lichtpaket zwar multipliziert statt addiert, aber deswegen geht sie doch mit einem roten Lichtpaket. von über 700 nm Wellenlänge, wenn auch sehr schlecht. Die Rotquanten sind viel zu schwach, um selbst das H+...OH- zu spalten. Die Wasserspaltung kann also auch schon durch das superschwache Leuchten ausgelöst werden, welches durch den Ultraschall hervorgerufene blaue Lichtblitze sein können (Sonolumineszenz). Die Wasserspaltung wird also auch nicht direkt durch das Sonnenlicht bewerkstelligt. Sonnenquanten sind notwendig aber nicht ausreichend, das Leben auf der Erde zu erhalten. Noch besser gesagt. Es ist nur die Energie der Sonnenstrahlung, die gebraucht wird. Pflanzen wachsen auch im Dunkeln mit Zuckerernährung. Wer spaltet dort den Wasserstoff von den Zuckern ab? Wer spaltet beim Citratzyklus das Wasser?
Hier möchten wir noch auf eine Arbeit (63) hinweisen, die eindeutig feststellt, daß bei allen äußeren Streßmöglichkeiten immer zuerst die Transpiration und eng gekoppelt damit der Ertrag abnimmt.
Große Hoffnungen legte man einst in die sogenannten Antitranspirantien. Das sind z.B. äußerst feine Folien, die auf die Blätter gesprüht werden, die Wasserabgabe der Pflanze drosseln, aber die Photosynthese kurzzeitig kaum beeinflussen. Man wollte so viel Wasser sparen, besonders in den Entwicklungsländern. Die Erträge nah-men mit abnehmender Transpiration natürlich proportional ab.
Das nächste schwierige Problem ist die Unterscheidung von unproduktiver Verdunstung und aktiver lebendiger Transpiration. Daß die letztere existiert zeigen besonders die Temperaturmessungen an Blattoberflächen. Wenn ich einen Wassertropfen auf die obere Blattfläche aufbringe, dann sinkt die Temperatur in unmittelbarer Umgebung des Blattes auf seiner Ober- und Unterseite GLEICHMÄßIG im Vergleich zu seiner Umgebung durch die Verdunstungskälte. Wenn ich die den Temperaturunterschied zwischen Umgebung und Blattoberfläche ohne Wassertropfen gleichzeitig auf der Blattober- und Unterseite messe, dann stelle ich sehr oft fest, daß beide eine um mehrere Grade unterschiedliche Temperaturdifferenz zur Umgebung haben. Die Versuche mit dem Wassertropfen zeigen, daß das Blatt kaum einen Leitungwiderstand für die Wärme darstellt, weil ständig die Wasserströme durchfließen und einen angeregten Wärmeaustausch erlauben. Bei einigen Gewächshauspflanzen (64) wurde festgestellt, daß bei Zurückziehen des Wärmeschirmes am Morgen die Temperatur der Blattoberfläche plötzlich um mehrere Grade anstieg. Der Lichtanstieg durch den Rückzug des Wärmeschirmes muß zu einer Transpirationsvergrößerung führen. Eine schwarze Scheibe zeigte eine ziemlich genaue Anpassung an den Temperaturgang des Gewächshauses. Es bleibt nur der Schluß, daß die Pflanze die Temperaturabnahme durch die Transpiration variieren kann, daß das nicht nur Verdunstungskälte ist. In (61) wurde auch gezeigt, daß Gurkenblattoberflächen auch durchgehend eine wärmere Temperatur als ihre Umgebung haben können. Wüstenpflanzen können bis zu 10 K unter die Umgebungstemperatur abkühlen, was niemals Verdunstungskälte sein kann. Die starke Abkühlung bei den Wüstenpflanzen ist wahrscheinlich ein Trick, um die Luft anzufeuchten, die in der Wüste sehr trocken ist. Trockene Gase können nicht von Lebewesen aufgenommen werden. Denken Sie an die Fenstergläser die beschlagen, wenn sie kälter als die Raumtemperatur sind.
Ein australischer Autor (65) maß über das ganze Jahr die Wasserabgabe eines Eucalyptusbestandes im Vergleich zur Wasserabgabe eines Wasserkessels. Der Eucalyptusbestand hatte eine größere Wasserabgabe. Trotzdem war der Temperaturabfall über dem Wasserkessel viel größer als durch den Eucalyptusbestand. Wenn ich einen Zweig mit Wasser besprühe, dann ist der Temperaturabfall durch die Verdunstungskälte genauso groß wie das durch die verdunstete Wassermenge auch sein muß. Wenn ich die Wassermenge, die der unbenetzte Zweig durch seine Transpiration freigibt, zugrundelege, dann sinkt seine Oberflächentemperatur nicht um die aus der Verdunstungskälte berechnete Menge. Um noch ein bißchen bei dem Eucalyptusbestand zu bleiben. Er transpirierte viel mehr Wasser als durch die Niederschläge runterging. Er mußte das Wasser über 100m hochpumpen und machte so viel Transpiration. Wie tief da die Wurzeln runtergehen müssen, um das Grundwasser zu erreichen.
Wenn ich eine Energiebilanz des Gewächshauses aufstelle muß ich die Verdampfungsenthalpie natürlich berücksichtigen. Ich kenne die Bodenverdunstung und die pflanzliche Transpiration, aber der Pflanzenbestand kühlt das Gewächshaus nicht um den Betrag der Wassermenge, die er transpiriert. Das wird noch unverständlicher, wenn man weiß, daß durch den Woodeffekt die wärmebringende Strahlung durch die Pflanzen reflektiert wird. Denken Sie an den kühlenden Waldesschatten.
Ich möchte in dem Zusammenhang an die gewaltigen Wälder Sibiriens erinnern, die trotz ihrer riesigen Ausdehnung und Riesenwassermassen, die sie jährlich vertranspirieren, kaum eine Abkühlung wie der Atlantische Ozean bringen. In Rußland herrscht Kontinentalklima, in Westeuropa Meeresklima. Die Pflanzen wandeln das sichtbare Licht in Wärme für die Transpiration und damit Lebensenergie. Das blanke Wasser verwandelt die infrarote Strahlung in Wärme für die Verdunstung. Pflanzen nehmen kein Infrarot auf (Woodeffekt). Die Wärmeenergie, die bei der Absorption der infraroten Strahlung im Boden oder woanders entsteht, nehmen sie natürlich gerne indirekt durch die Wasseroberflächenaufnahme zu sich. Auch Blätter nehmen bekanntlich Wasser auf. Es gibt im Gartenbau die sogenannte Blattdüngung. Das H+...OH- kann direkt durch die Epidermis dringen, weil es hydrophob ist. Auch die Wurzelspitzen sind hydrophob gepanzert und außerdem noch luftdicht. Die Pflanzen nutzen dadurch irgendwie das gesamte Sonnenspektrum, ohne viel Energie durch Verdunstungskälte zu verlieren. Aktive Wasserabgabe der Zelle ist weit ökonomischer als Verdunstung oder Verdampfung, weil sie nur T-Wasser abgeben. Sie nehmen nur Wasseroberflächen auf und geben sie später wieder ab als T-Wasser. Das ist der Riesenvorteil der Oberflächenwesen. Es ist keine Verdunstung sondern vor allem eine SUBLIMATION. Denken Sie an die lebendigen Eisstrukturen. Ein Tripelpunkt bei starkem Druck in den Mikrostrukturen (Laplacesches Gesetz) ist um die 40 Grad Celsius.
Vielleicht sind darum auch die homoiothermen Vögel und Säuger so unökonomisch, weil sie das T-Wasser, das die Zellen abgeben, durch die Körperwärme gleich wieder in natürliches Wasser (Schweiß) verwandeln müssen, was viel Energie kostet. Vögel nennt man Sklaven der Nahrung. Der Mensch ist der größte Verschwender. Der alte Römer brauchte ständig eine Pferdestärke im Laufe eines Jahres, wir brauchen wieviel?
Übrigens haben Insekten auch um ihre Zellen viel Luft genau wie die Blätter (Interzellularen und die Tracheen), so daß sie sehr gut Sublimation machen können. Die Tracheen bringen die Luft direkt bis in die Zellen.Viele sekundäre Wasserpflanzen haben ein Aerenchym, so daß sie sublimieren können. Der Phasenübergang vom Eis in Gas ist wesentlich energiegünstiger als von Eis in Flüssigkeit. Der Nachteil ist bei so viel Interzellularen die ungenügende Festigkeit. Die Insekten haben deshalb wohl das Außenskelett entwickelt, um sich die energiegünstigen Interzellularen leisten zu können. Dafür konnten sie nicht so groß und mächtig wie die Wirbeltiere mit dem Innenskelett werden, die den Menschen mit dem großen Gehirn hervorbrachten.
Das die Transpiration vor allem ein aktiver sinnesphysiologisch beeinflußter Prozeß ist, zeigen folgende Versuche.
1. Die Pflanze verlernt die Transpiration, wenn sie längere Zeit im Dunkeln steht. Auch nach Zurückstellen ins Licht braucht sie lange Zeit, um wieder auf Transpirationsraten zu kommen, die sie vor der länger andauernden Verdunklung zeigte (28).
2. Wenn ich in einer Klimakammer die Transpiration messe und dann etwas sogenanntes far red Licht dazugebe, das keine energetische Komponente darstellt, weil es wirklich nur in geringsten Mengen deutlich wirkt, so steigt die Transpirationsrate unverhältnismäßig stark an. Alle Lebewesen erleben die Umwelt nur vermittelt über aktiv erzeugte Sinnesenergie (Müllergesetz). Man kann sie deshalb leicht betrügen, wenn man die Kopplung, die in der Natur zwischen Sonneneinstrahlung und far red herrscht, aufhebt. Warum wirkt das far red so stark? Dieses far red ist dasselbe für die Pflanze, was für uns das sichtbare Licht ist. Ihre tausenden von far red-em-pfindlichen Stellen arbeiten vor allem um die Mitpflanzen zu sehen.Je mehr Mitpflanzen, um so stärker muß sie wachsen, um im Licht zu bleiben.
3. Wenn ich Gurkenpflanzen im Gewächshaus durch ständiges Geschlossenhalten der Lüftungklappen einer hohen Luftfeuchte aussetze, verstärkt sie ihren inneren Druck zum Antrieb der Transpiration so (66), daß nach Öffnen der Lüftungsklappen, wodurch das Sättigungsdefizit der Luft zunimmt, die Wasserabgabe so stark steigt, daß die Blätter an einigen Stellen austrocknen und ganz weiß werden. Die Pflanzen erleiden einen Schock und erholen sich oft nicht mehr davon. Die Transpiration nimmt auf die Dauer ab.
Wenn ich ein mathematisch-statistisches Modell der Transpirationsmenge oder -rate in Abhängigkeit vom Licht und Sättigungsdefizit mache, dann erreiche ich eine sehr gute Anpassung des Modells an die Wirklichkeit. Wenn ich aber die Luftfeuchte mit hinein nehme als Extragröße, so kommt paradoxerweise heraus, daß mit steigender Luftfeuchte eine stärkere Transpiration stattfindet, obwohl ins Sättigungsdefizit die Luftfeuchte mit eingeht. Die Luftfeuchte hat also keinen rein physikalischen Einfluß über die Saugeigenschaften der Luft. Der Wassertransport wird nicht direkt durch das Sättigungsdefizit der Luft getrieben.
Bei sehr geringer relativer Luftfeuchte um 30% ist die Transpiration sehr stark, aber der Ertrag äußerst niedrig (67). Dann kann die Pflanze nicht mehr oder kaum noch aktiv transpirieren. Die unproduktive Verdunstung triumphiert.
Auch das lebendige L-"Wasser" wird der Zelle direkt entrissen, ohne sich in T-Wasser zu wandeln.
Die Pflanzen brauchen auch einen gewissen Widerstand, eine Schwelle, um den aktiven Transpirationsdruck aufzubauen.
Leider konnte ich diese Transpirationstheorie nicht bis ins notwendige Detail ausarbeiten, weil man mich aus Großbeeren, wo ich jahrzehntelang beschäftigt war und wo ich mein Ertragsmodell erarbeitet habe, aus politischen Ungehorsamkeitsgründen rausgegrault hat. So konnte ich auch nicht den ungemein wichtigen Wasserfaktor ins Modell einarbeiten, sondern mußte von der Voraussetzung ausgehen, daß das Wasser immer im Optimum zur Verfügung steht. Bis jetzt hat sich auch keiner bereiterklärt, mir bei der Ausarbeitung behilflich zu sein. Ich denke aber, daß die angeführten Tatsachen eine gute Stütze für meine Lebenstheorie sind, wenn auch für die Ausnutzung der Erkenntnisse in der Praxis noch einiges getan werden muß. Das eigentliche Hohelied der Aktiven Transpiration (AT) wird jetzt gesungen, wenn wir die Schließ- und Nebenzellen des Spaltöffnungsapparates betrachten. Das sind die Weltmeister der AT. Angeblich dienen die Spaltöffnungen zum Schutz vor zu starker Austrocknung, weil sich bei Wasserstreß die Öffnungen schließen. Das ist aber auf Dauer gesehen falsch. Versuche in klimatisierten Küvetten mit ganzen Pflanzen darin zeigen, daß eine sehr schnelle Änderung des Lichtes oder der Temperatur zu einer sofortigen Reaktion der Transpiration führen. Die Öffnungsweite der Spaltöffnungen kann niemals so schnell folgen, wie nachgewiesen wurde. Die Spaltöffnungen folgen der Transpiration hinterher und steuern sie kaum. Es ist also genau umgekehrt, wie es in vielen Büchern steht. Für die Öffnung ist die noch zu diskutierende peristomatäre Transpiration zuständig, die die direkt durch die Schließzellen oder in der Nacht durch die Nebenzellen führt. |
|
|
|
|
|
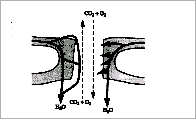
Abbildung 12 Stark schematisierter Querschnitt durch eine Spaltöffnung im Blatt mit Hauptgaswegen
|
|
|
|
|
|
Je wärmer und trockener das angestammte Biotop einer Pflanzenart ist, um so höher ist die Spaltöffnungszahl pro Blattflächeneinheit. Wüstenxerophyten haben 129, Wasser- und Sumpfpflanzen 61, Pflanzen des tiefsten Waldesschattens nur 46 pro Quadratmillimeter. Letztere haben den geringsten Wasserstreß, weil sie kaum Licht empfangen. Wenn ich Blätter Wasserstreß aussetze, schließen sie natürlich zuerst die Spaltöffnungen. Je länger aber der Wasserstreß andauert (Tage, Wochen, Monate), um so größer muß ich das Wasserdefizit des Bodens steigern, damit sich die Spaltöffnungen schließen.Blätter der gleichen Klone an feuchten Standorten haben ebenfalls wie die angestammten Schattenpflanzen wesentlich weniger Spaltöffnungen pro Flächeneinheit als die Blätter der am trockenen Standort stehenden Pflanzen.Die Aufgabe ist es also, mehr eine größere Transpirationsbereitschaft aufzubauen- die Transpiration um jeden Preis auf die Dauer aufrechtzuerhalten. Sie haben eine integrierende Funktion. Außerdem sollen sie eine indirekte informatorische Kopplung zwischen Transpiration und Gasaustausch des Sauerstoffs und Kohlendioxids gewährleisten, weil die AT die Photosynthese und die Atmung antreibt. Beide Prozesse sind auf diese Gase angewiesen. Auch hier hat also die AT das Primat.
Das ist alles mehr oder weniger wahr, jedenfalls wahrer als die alten Theorien. Richtige Klarheit erhielt ich aber erst, als ich den Atmungsstoffwechsel der Lunge näher angesehen hatte, worauf mich ein alter, erfahrener, medizinischer Assistent aufmerksam machte. Die Lunge kann nur richtig atmen, wenn die Atmungsgase angefeuchtet werden. So kam ich dann auf den Trichter, daß auch die Spaltöffnungen die Hauptaufgabe haben, die Gase anzufeuchten, damit sie von der Zelle aufgenommen werden können. Kohlendioxid und Wasser sind der Komplex, der bei der Photosynthese vereinigt wird. Wenn die Luft sehr trocken ist , dann müssen mehr Arbeiter pro Flächeneinheit vorhanden sein, um die Kontakthäufigkeit zwischen Luft und wasserspendender Fläche zu erhöhen. Die Schließzellen können nur unter starkem Druck, im geöffneten Zustand also, richtig Wasser in die vorbeiströmende Luft einsprühen, so daß es leicht erklärlich wird, daß sich nach längeren Dürreperioden die Spalt-öffnungen bei dem gleichen Wassermangel später schließen als in den guten Zeiten der ausreichenden Wasserversorgung. Wüstenpflanzen greifen dann sogar zur Kühlung der Luft, um die relative Feuchte so weit wie möglich zu erhöhen.
Im geöffneten Zustand der Spaltöffnungen, bei gespannten, aufgepumpten Schließzellen, arbeitet die Pflanze am stärksten. Das Gros des Wassers geht dabei durch die Schließzellen selber. Des Nachts geht das meiste durch die Nebenzellen. Die Wasserabgabe durch die Schließ- und Nebenzellen ist voll bewiesen. Hier wird nur gesagt, daß die HAUPTmenge durch sie hindurchgeht. Die letzte Theorie stammt von Herrn Stahl, der die einschlägigen Experimente schon im vorigen Jahrhundert machte (68). Hier einige experimentelle Ergebnisse:
1. Man stellt Blätter mit dem Stiel in schwermetallsalzhaltige Lösungen (Thallium, Silber), läßt sie transpirieren im Licht und so wandern die Schwermetallsalze mit dem Transpirationsstrom und bleiben natürlich dort liegen, wo sich das Wasser in Gas verwandelt (70). Das ist eine BIOAKKUMULATION, das lebendige Analogon zur Destillation, die wir schon besprochen haben. Fast alle Schwermetalle finden sich am Tage in den Schließzellen und des nachts in den Nebenzellen!!! Kommentar eigentlich überflüssig, aber in diesem Falle trotzdem nicht, denn ich habe mich an Frau Maier (70), die diese Experimente mit Akribie aber ohne Phantasie wiederholte, gewandt und meine Theorie kurz dargestellt, aber sie hat meine Argumente abgelehnt, weil sich beim Löschpapier auch Farbstoffe am Rande sammeln. Das sei nur ein Randeffekt das mit den Schwermetallen. Ich kann vorerst, ehe ich zu weiteren Beweisen komme, nur sagen, daß der Randeffekt nur dort zustande kommen kann, weil die Berührungsfläche mit der Luft am Rande natürlich am größten ist und folglich auch die Wasserabgabe am stärksten ist. Ob ich das nun Randeffekt nenne oder nicht, die Wasserabgabe ist direkt von der Oberfläche abhängig. Frau Maier hat natürlich recht, daß dieses Experiment alleine nicht ausreicht.
2. Das zweite Experiment zeigt den überaus starken Druck unter dem die Schließzellen stehen. Wenn ich mit einem Mikromanipulator in die Schließzellenwand einsteche, verkleinern sie sich sofort auf ihre Nachtgröße. Sie pumpen sich nicht durch osmotischen Wasserdruck auf sondern blasen sich durch T-Wassergasabgabe auf.
3. Sie kühlen sich trotz der großen Wasserabgabe nicht ab. Sie bedecken nur 1% der Blattoberfläche und machen zumindestens peristomatäre Transpiration. Folglich führt das Wasser die Energie zur Transpiration heran, es ist kaum Luftwärme gefragt in Form von Verdunstungs- kälte.
4. Bei reiner Wasserabgabe mittels Verdunstung durch die Mitte der Spaltöffnungen könnte kaum ein CO2-Gegenstrom zur Photosynthese passieren und die Photosynthese müßte bei steigender Wasserabgabe sinken. Genau das Gegenteil ist der Fall. Transpiration und Photosynthese steigen fast immer proportional.
5. Bei einem amphistomatären Maisblatt blies ein Forscher (69) einen Luftgegenstrom durch die Stomata. Er konnte die Transpiration nur um die Hälfte absenken, weil er nicht den oberen Teil der Schließ- und Nebenzellen erreichte.
6. Nachts sammeln sich die Schwermetalle in den Nebenzellen, warum tritt da nicht der Randeffekt in Erscheinung Frau Maier? Ich kann die Sache erklären, denn nachts schließen sich die Öffnungen und hinderten so durch einen Transpirationsgasstrom die Sauerstoffdiffusion zur Atmung stark. Aber die kluge Pflanze verlegt die Transpiration in die Nebenzellen.
7. Ein anatomisches Argument gibt es auch. Die meisten Ektodesmen der Blattoberfläche gehen zu den Spaltöffnungen.
8. Man verschloß durch einen Trick die Spaltöffnungen mit Wachs, aber genau wie beim Entgegenblasen nahm die Transpiration bei Blättern und Früchten kaum ab (71, 72). Kutikuläre Transpiration kommt kaum in Frage, denn schon Praktikumsversuche zeigen, daß Blattseiten ohne Spaltöffnungen kaum Transpiration zeigen. Die stark wasserabweisenden Wachsschichten auf den Blättern und Früchten sind so dick, daß da kaum Wasser durchkommt. Es ist ja auch ihre Aufgabe, den Weg der Wassermassen zu den Spaltöffnungen zu erzwingen. Schauen Sie auf ein regennasses Blatt, wie das die Wassertropfen veranlaßt sich zusammenzukugeln und abzurollen. Schauen Sie in das Mikroskop, wo es durch Anfärben gelungen ist zu zeigen, daß fast alles Wasser zu den Spaltöffnungen strömt.
9. Spaltöffnungen hindern nicht die Transpiration, denn Tomaten- und Gurkenpflanzen transpirierten mit fast unveränderter Geschwindigkeit bis zum Tode durch Austrocknen. Transpiration ist wichtiger als Verhinderung und Wasser sparen.
10. Es gibt außer den Wachsverschlüssen und Gegenblasen noch einen sehr schönen Beweis für die getrennten Wege von Wasser und den Atmungsgasen. Wenn ich nämlich ABA gebe, dann schließen sich fast immer, außer bei Kälte z.B., die Spaltöffnungen. Die Transpiration nimmt aber wesentlich stärker als die Photosynthese ab. Bei einer Pflanzenart nahm die Photosynthese sogar zu. ABA setzt die Wasseranziehungskraft der Schließzellen stark herab. Die Öffnungsweite spielt als Diffusionshindernis keine so große Rolle.
Der Quotient aus Transpirations- und Photosyntheserate hat bei der gleichen Art unter verschiedenen Klimabedingungen eine andere Größe. Bei der gleichen Pflanze ändert er sich in Abhängigkeit von den äußeren Bedingungen (76). Die Öffnungen sind nur dazu da, um die Kontaktfläche der Schließzellen mit der Luft zu erhöhen. So schafft die Pflanze auch gleichzeitig einen bequemen Eingang für die Atmungsgase. Sie macht auf Randeffekt! Nachts braucht sie den nicht, da ist die Anspannung wesentlich kleiner. Wenn ich aber nachts die Temperatur stark er-höhe, dann steigt der Streß und die Paranekrose und die Pflanze öffnet durch die Transpiration die Spaltöffnun-gen und erhöht den Randeffekt, aber auch die Öffnungsweite für die stärker anfallenden Gase. Sauerstoff rein zur Atmung, Kohlendioxid raus, weil nachts kaum Photosynthese ist. Wachstum und Luftanfeuchtung durch Transpiration um jeden Preis und wenn die unwichtigen Teile schrum-pfen.
11. Noch ein Experiment von einem meiner größten Gegner, der es bis heute ablehnt, mit mir überhaupt zusammen zu treffen, der die Osmose über alles liebt und so natürlich gegen mich ist (73). Er beweist schön die These, daß Wachstum und Transpiration und somit Öffnungsweite der Stomata genau dieselben Seiten einer Medaille sind. Alles, was das Wachstum fördert wie Wachstumshormone, Kalium, Licht, Temperatur (Wärme) öffnet die Spaltöffnungen. Die Transpiration wird ebenfalls durch diese Faktoren gefördert. Die jetzige Theorie geht davon aus, daß Kalium in die Spaltöffnungen gebracht wird, das den osmotischen Wert stark steigert und somit Wasser anzieht. Wenn man aber Gibberellin oder Kinetin, zwei sehr stark wachstumsfördernde Hormone, gibt auch nachts, so öffnen sich die Spaltöffnungen weit ohne Kaliumein- strom!!!
Kalium und Calcium aber auch Magnesium spielen beim Regeln des Wasserzustandes des Plasmas eine entscheidende Rolle. Sie sind auf die Dauer auch durch Hormone nicht zu ersetzen. Die Pflanze läßt sich nicht für alle Zeiten betrügen.
Ein noch schöneres Experiment machten andere Forscher (74). Sie stellten dabei fest, daß Inhibitoren, die die Bildung der alterungsfördernden Proteine hemmen (programmierter Tod), die Spaltöffnungen vergrößern. Inhibitoren, die nur allgemein die Eiweißsynthese hemmen, sowohl die der alterungsfördernden als auch die der gegenteiligen, haben keinen Einfluß auf die Spalt- öffnungsweite.
12. Noch zwei Beweise für die relativ unbedeutende Rolle des Schlitzes im Spaltöffnungsapparat. Auch junge Blätter, die noch keine Spalten im Stomataapparat haben, zeigen eine lebhafte Transpiration. Auch bei Wüstenpflanzen, deren Blätter dick gepanzert sind. Man sollte mal die Nadeln von Kiefern untersuchen, denn Nadelbäume sind oft Xerophyten mit geringem Wasseraustausch und dicken Wachsabdeckungen gegen unfreiwillige Verdunstung.
Wenn ich Blätter mit Turgorin behandle (75), so schließen sich die Spaltöffnungen, aber die Transpiration nimmt nicht einen Deut ab. Vielleicht haben die Spaltöffnungen unterschiedliche wasserdurchlässige Bezirke auf ihrer dicken Zellwand, die dem starken Druck bei der T-Wasserabgabe widerstehen muß. Einige lassen mehr Wasser nach oben, andere mehr nach innen ab.
Auch die homoiothermen Vögel und Säuger haben den stärkeren Wasserdurchsatz als die Reptilien und Amphibien. Außer bei genügend Außenwärme, wenn die letzteren poikilothermen Tiere sehr agil werden können. Letztere sind dafür wesentlich sparsamer und ökonomischer als die so aktiven warmen.
Wenn ich Pflanzen mit Kohlendioxid im Gewächshaus begase, dann steigt die Trockenmasse nur um rund 30%, aber die Photosynthesegeschwindigkeit um über das zwei- bis dreifache.
Wachstum ist in erster Linie L-"Wasser"einbau und nicht Trockenmassezunahme, auf die der Mensch natürlich den größten Wert legt und auch alles auf sie bezieht und sie sogar Biomasse nennt.
Durch Messungen an Blattscheiben konnte ich eindeutig zeigen, daß bei der Photosynthese Trocken- und Frischmasse steigen.
Die Photosynthese ist keine reine Stärke-, Zucker- oder ATP-Zunahme. Sie ist vor allem Wassereinbau - eine Frischmassezunahme.
Bei der Betrachtung der Spaltöffnungen muß ich immer an das Müllergesetz denken. So viele Einflüsse, eine Allerrregbarkeit, aber sie reagieren nur stereotyp mit Wasserabgabeverstärkung oder - verminderung.
Das kann man auch mit dem Blutdruck verglei-
chen, der allerregbar ist, aber nur steigen oder sinken kann. Das Herz ist ein sinnesphysiologisches sehr altes Zentrum. Die Spaltöffnungen sind Herzen der nicht zentralisierten Pflanzen. Ich möchte hier nur auf eine Arbeit verweisen (77), welche die überragende Rolle des Calciums für die sinnesphysiologischen Spaltöffnungen zeigt. Ohne Calcium verfallen sie in einen Auto rhythmus. Sie bleiben dauergeöffnet.
Damit wäre ich am Ende des biologischen Teiles des Buches. Ich konnte fast alles nur andeuten, denn Du sollst Dir kein Bild machen und lieber denken. So bin ich auch sehr sparsam mit Abbildungen. Auf das Wesentliche im Leben, auf das, was alle Lebewesen gemeinsam haben, wollte ich Sie hinweisen. Auf die Kontaktflächen, die Wärme- und Liebebedürftigkeit und vor allem auf das hydrophobe, wasserfeindliche H2O.
Zum Schluß kann ich nicht umhin, wieder den alten Einstein zu kitzeln. Er hat den Nobelpreis nicht für seine Relativitätstheorie bekommen, die wir auch schon wesentlich vereinfacht haben durch die Annahme der unendlichen Nullgeschwindigkeit. Letztere Theorie könnte auch besser erklären, warum in Golddrähten eine wesentlich höhere Lichtgeschwindigkeit herrscht als im Vakuum, wo es doch unübertroffen schnell sein soll ( 300 000 km/sec.). Er postulierte die Lichtquanten, die Photonen. Damit erklärte er u.a. die mechanischen Eigenschaften des Lichtes, das einen Druck ausüben kann. Die Photonen prallen auf ein Schaufelrad und treiben es an.
Durch das Auftreffen der Lichtwellen auf ein Schaufelrad wird die Materie erwärmt, kann sich aber nicht fortbewegen, weil Gase kaum Wärme leiten Bei Wärme dehnen sich alle Körper aus, weil ihre Atome und Mo-leküle mit einer größeren Amplitude schwingen, so daß das Licht letztendlich in mechanische Energie verwandelt wird - das Schaufelrad beginnt sich zu drehen. Dafür gibt es auch ein mathematisch-physikalisch formuliertes Umwandlungsgesetz, was ich als dummer Biologe natürlich nicht weiß.
Wie ich zu dieser ungeheuerlichen Behauptung gekommen bin? Meine zweitbeste Freundin Marlene und auch viele andere Menschen haben ein ähnliches Flügelrad in ihrem Wohnzimmer zu stehen.
Meistens am Fenster, weil Herr Einstein das Licht so sehr liebte. Nur hat dieses Rad eine Besonderheit, denn eine Seite der Schaufel ist schwarz und die andere Seite viel heller. Das ist bei allen anderen Schaufeln, die auf demselben Rade befestigt sind, ebenfalls so. Schwarz kann am besten Lichtenergie in Schwingungswärme verwandeln.
Das Rad dürfte sich eigentlich nicht drehen, wenn Einstein recht hätte, denn Photon ist Photon, egal auf eine wie auch immer gefärbte Fläche es trifft. Ich habe das so gelernt, aber die Verkomplizierer werden schon eine Widerlegung finden in der Einmaligkeit eines Genies wie Einstein, der nicht einmal die amerikanische Sprache richtig beherrschte, obwohl er lange dort lebte. Ich habe einen sterblichen Gott gestürzt. Glücklich bin ich deshalb nicht. Stürzen ist gemein und häßlich und man beleidigt unzählige Fans oder Jünger. Ein Grund mehr, dieses Buch nicht zu drucken.
Man müßte einmal die beiden Seiten aller Schaufeln mit verschiedenen Farben bemalen und auch eine andere Oberflächenstruktur geben und zum Wettbewerb aufrufen, wer das schnellste Rad baut, bei dem aber nur die Oberfläche der Schaufeln verändert werden dürfte. Alles andere muß standardisiert werden. Ich stifte den Augustinschen Licht- radpreis. 1000 Mark könnte ich bieten. Es würde leider viel teurer werden. Das Lichtrad meiner geliebten Marlene könnte den Standard in seinen Abmessungen abgeben. Auch die Gasumgebung dieser Lichtmühle müßte genormt werden, eben alles außer den reinen Oberflächen. Man könnte die Theorie der Photosynthese revolutionieren.
Diese Lichtmühle hat mich dazu inspiriert, die Rolle des Carotins besser zu verstehen. Sehen Sie sich einmal das Carotin an. Es hat im Molekül eine lange Kette, die an zwei aromatischen Verbindungsringen aufgehängt sind. Denken Sie sich die schwarz-weißen Flächen der Lichträder auf eine Achse aufgereiht wie die Moleküle in der Kette des Carotins. Denken Sie an eine der Funktionen der Carotine, die sie besonders bei starkem stressigen Licht haben. Sie können Wärme abführen, indem sie diese wahrscheinlich in Rotationsenergie und damit in mechanische Kraft für die Photosynthesefermente wandeln.
Neutrinos, Photonen u.s.w. sind nur mehr oder weniger geschickt gefangene Wärmeströmungswirbel. An Oberflächennetzen geht das am besten. Auch in tiefen Bergwerken mit den sogenannten Neutrinofallen, was weiter nichts ist als eine riesige Wassermasse mit Cerenkovzählern, können sich immer wieder mal einige Wärmeströme an einem Wassermolekül versammeln und es zum Blitzen bringen. Es gibt nicht nur Alfa-, Beta und Gammastrahlung - es werden ganze Atome abgestrahlt, wie das in den
letzten Jahren nachgewiesen wurde. Der Zufall läßt das immer wieder zu. Man weiß nur daß, aber nie wann und wo genau.
Ein Proton ist ein fast ganzes Wasserstoffatom - nur ohne Elektronen ..... Denken Sie nicht wie ich in diesem Moment auch an Transmutation??? |
|
|
Druckbare Version
|
|
|
3.2. Das Wachstum
|
|
|
|
|

